UCLA Department of Chemistry and Biochemistry
153AH - Fall 2009 - Instructors: Todd Yeates, Duilio Cascio, Tobias Sayre
Human Topoisomerase I
by Albert Aparicio
|
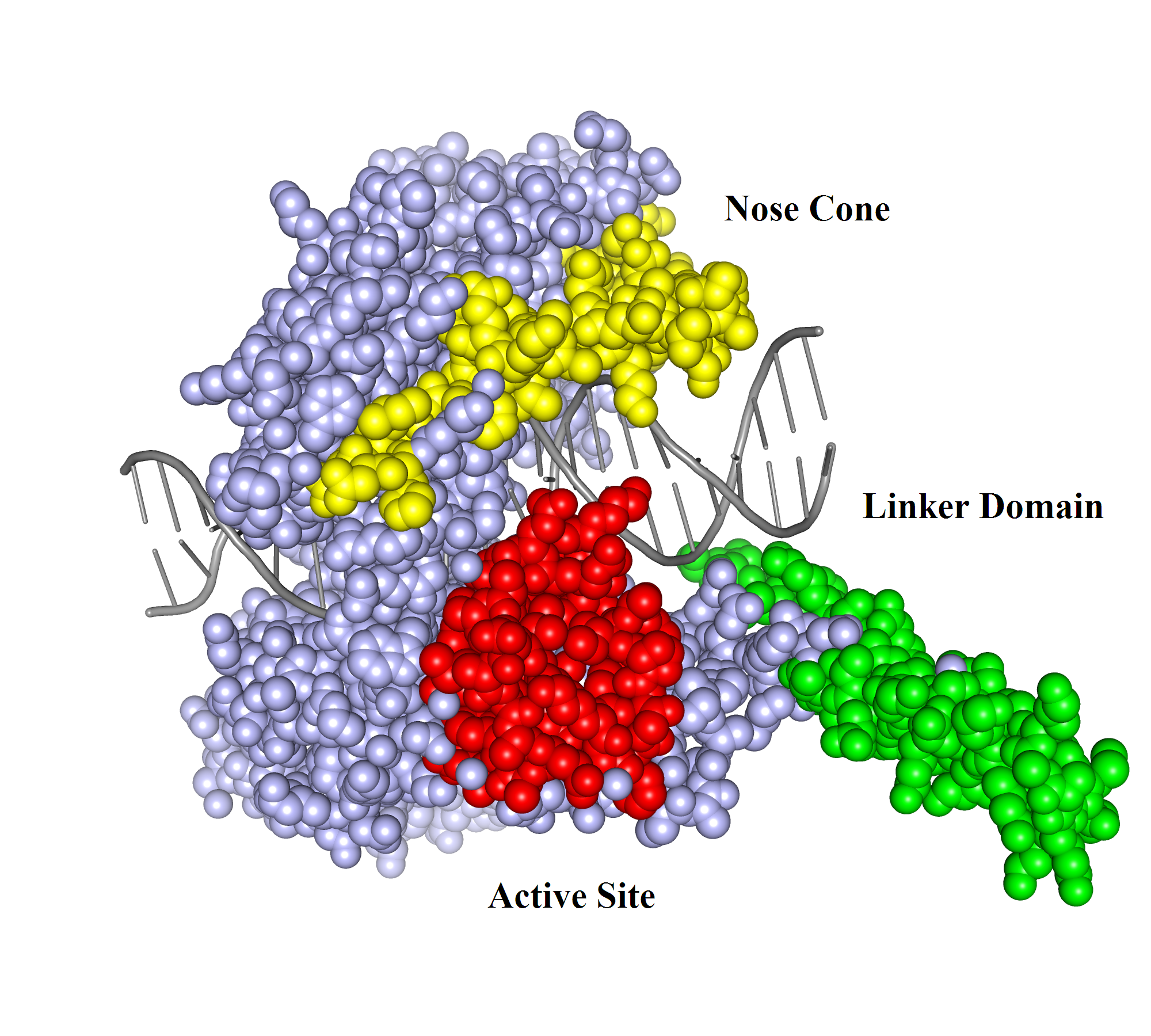
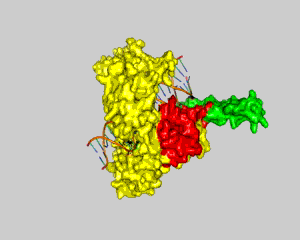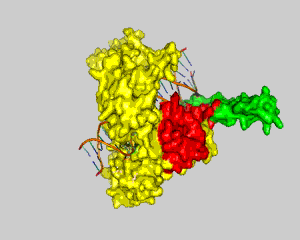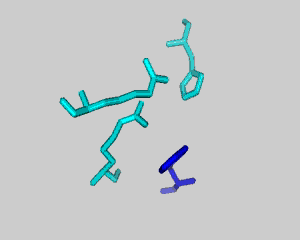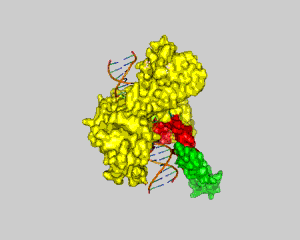
DNA can efficiently compress itself through supercoiling of its double-stranded helix structure. Nevertheless, this geometric shape can hinder cellular processes such as transcription and DNA replication. However, there are enzymes called topoisomerases that can solve these problems through unwinding and untangling of the negative and positive supercoiling in DNA. Two types of topoisomerases are known: topoisomerase type I and topoisomerase type II. Topoisomerase type I reduces tension in supercoiled DNA by cleaving one strand of the double-stranded helix and allowing it to unwind itself until religation. Topoisomerase type II has the same purpose, but cuts both strands of a DNA and allows passage of another segment of double-stranded DNA helix through the cleavage site. Topoisomerase type I can be subdivided into two subcategories: type IA and type IB. Type 1A is categorized by covalent binding of the enzyme to the 5′ end of the broken DNA strand and type IB is categorized by covalent attachment of the enzyme to the 3′ end (1). An example of a topoisomerase type IB is Human Topoisomerase I. Human Topoisomerase I is a 765-aa enzyme that clamps around the DNA like a clam. It contains four discrete domains (Fig. 1). The N-terminal domain is made up of residues 1-214. It has not been ascribed any significance to the enzyme's function and is poorly conserved (1). Adjacent to the N-terminal domain is the Core domain (residues 215-634). It is highly conserved and consists of subdomain I, subdomain II, and subdomain III. Subdomain I and II make up the upper lobe of the enzyme called the capping lobe. The last two domains are the Linker domain (residues 635-696) and the C-terminal domain (residues 697-765). These two domains, along with subdomain III, form the lower lobe called the catalytic lobe. The two lobes are hinged together by the eighth alpha helix on one side of the DNA, and the tips of the lobes form a "lips" region on the other side of the DNA helix. The formation of the lips region is due to salt bridge interactions (1). DNA relaxation by Human Topoisomerase I begins with a nucleophilic attack by tyrosine (Y723) located in the active site (residues 723-765) in the C-terminal domain. It introduces a transient break into one strand of the DNA and forms a covalent intermediate with the 3′ end of that strand (2). There are also three residues in the core domain aiding in stabilization of the phosphate backbone following cleavage by the catalytic tyrosine. Two arginines, R488 and R590, form hydrogen bonds with the oxygen atoms of the scissile phosphate (2). Histidine (H632) also aids in stabilization of the scissile phosphate via hydrogen bonding, but might also function as a general acid. It possibly donates a proton to the 5′ end of the broken DNA strand (leaving group) during the cleavage reaction (2). Once cleaved, the unbounded broken strand untwists itself downstream of the cleavage site. This change of topology does not require energy of any kind because the untwisting of the helix is favored by the direction of the torsional strain (2). There are two other major regions of the enzyme, besides the active site, that guide the broken strand through DNA relaxation: the Nose Cone (residues 303-345) and the Linker domain (Fig. 2). They are highly conserved in positively charged amino acids and play an important role in topoisomerization. When the broken strand unwinds, it comes in contact with the DNA-proximal surfaces of both the nose cone helices and the linker domain (2). These positively charged surfaces match with the negatively charged DNA backbone and help regulate the winding process through ionic interactions. This process is called controlled rotation. When the DNA strand is relaxed through one or more turn cycles, the 5′ end of the broken strand comes back to the active site and re-ligates with the 3′ end of the DNA. This is done tyrosine acting as a leaving group while the 5′ end attacks the phosphate group of the DNA backbone. Thus the enzyme is removed from the DNA strand and can subsequently proceeed to the next DNA strand for another cycle of DNA relaxation. References (1) Carey, et al. (2003). DNA relaxation by human topoisomerase I occurs in the closed clamp conformation of the protein. Proc. Natl. Acad. Sci. USA 100, 5640-5645. (2) Stewart, et al. (1998). A Model for the Mechanism of the Human Topoisomerase I. Science. 279, 1534-1541. (3) PDBID 1a36 |
|