UCLA Department of Chemistry and Biochemistry
153AH - Fall 2009 - Instructors: Todd Yeates, Duilio Cascio, Tobias Sayre
Ops*-GαCT: Opsin in its G-Protein-Interacting Conformation
by Siyun Teresa Zheng
|
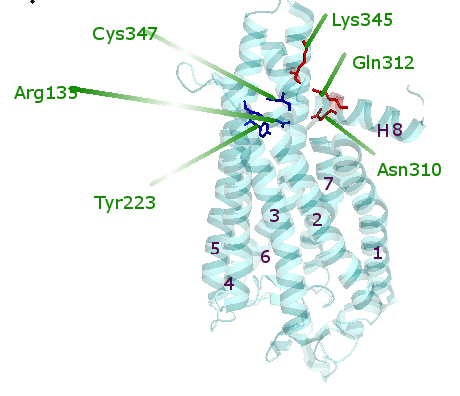
In the retina of the eye, the protein rhodopsin responds to dim light in the membrane of the rod photoreceptor cell. To date, rhodopsin is one of the best studied G-protein-coupled-receptors (GPCR). Rhodopsin is composed of a protein and a prosthetic group. The apoprotein is referred to as opsin. The prosthetic group is 11-cis retinal. The latter group is a light-absorbing pigment molecule that determines the color of rhodopsin and its responsiveness to light. Under illumination, the 11-cis retinal chromophore isomerizes to an all-trans geometry after photon absorption. This leads to the formation of the active, G-protein-binding metarhodopsin II state. Metarhodopsin decays when all-trans retinal is photolyzed and the protonated retinylidene Schiff base is hydrolyzed from its binding pocket; a ligand-free opsin is generated. In a low pH buffer, opsin is converted to its active G-protein-binding state, known as Ops*. Ops* is stabilized by an 11-mer oligopeptide from the guanine nucleotide-binding protein Gαt subunit, in what is referred to as the Ops*-complex (fig. 1). This product resulting from the loss of all-trans-retinal from the metarhodpsin II-GαCT complex, is considered to give an explanation for signal transfer from the receptor to the G-protein binding site (1, 2). Ops* has seven transmembrane helices connected by extracellular and cytoplasmic loops, and the cytoplasmic helix8. The hydrophobic residues in TM 5 and 6 in Ops* form a hydrophobic surface for the interaction with GαCT. When Ops* is induced by GαCT binding (fig. 2), the Ops*-GαCT complex shows changes in its protein interactions. The Ops*-GαCT complex has 326 residues in opsin, but 22 of these residues are not observed, and are believed to have high mobility. Opsin in its G-protein-interacting conformation is shown in figure 1, with the Ops*-GαCT complex lying along the membrane. Each of the Ops*-GαCT molecules is perpendicular to the intercellular and the extracellular surfaces. Ops*-GαCT shows electron density for Lys 296 in the retinal attachment site, but electron density is not observed in the retinal attachment site of Ops*. This indicates that GαCT binding stabilizes the Lys 296 in a potential network of weak interactions involving LysĀ296 in TM7, TyrĀ268 in TM6, and SerĀ186 and GluĀ181 in loop E2. In one case, GαCT homologues cannot bind to Ops* if a mutation occurs in the TM7-H8 kink (fig. 3) (1). A single receptor interacts with various G proteins and then triggers diverse signaling pathways (2). The structure of Ops*-GαCT provides clues about how the signal for GDP-GTP exchange is transferred from the receptor to the G-protein. First, the Gαt C-terminal α5 helix is modified to bind in the Ops*-GαCT complex. With the help of the α5 helix, GDP is released during the Ops* and GαCT interaction. In this case, the α5 helix acts as a transmission rod, and forces G-protein to be fixed for holding the receptor (1). On the other hand, whether single G-protein activates signal diversification is still controversial (2). References (1)Scheerer P., et al. (2008). Crystal structure of opsin in its G-Protein-interacting conformation. Nature 455, 497-502. (2)Tiina P. Iismaa, et al. (1995). G-protein-coupled receptors. Molecular Biology Intelligence Unit, 80-84. (3) PDBID 3DQB |
|